Библиотека
Теология
Конфессии
Иностранные языки
Другие проекты
|
Ваш комментарий о книге
Бондарев В. Концепции современного естествознания
Глава 9 ФЕНОМЕН ЖИЗНИ
§ 9.1. Сущность и уровни организации жизни
Сущность жизни и свойства живых организмов
На фоне возникновения и развития косной материи возникло и стало развиваться такое удивительное явление, как жизнь. В настоящее время описано более 1 млн видов животных, около 0,5 млн видов растений, сотни тысяч видов грибов, более 3 тыс. видов бактерий. Подсчитано, что не менее 1 млн видов пока остаются неописанными. Вопрос о сущности и происхождении жизни всегда имел для человека не только познавательный интерес, но и огромное значение для формирования мировоззрения. Возникновение и развитие живых существ вплоть до появления такого феномена, как человек, - одна из центральных проблем естествознания.
Живые организмы существенно отличаются от неживых систем. Эти отличия придают жизни качественно новые свойства. Рассмотрим сущность понятия «жизнь». Многочисленные формулировки сущности жизни можно свести к двум основным: 1) жизнь определяется субстратом, носителем свойств (например, белком); 2) ее трактуют как совокупность специфических физико-химических процессов. Например, Аристотель определял жизнь как питание, рост и одряхление; Г. Тревиранус - как стойкое единообразие процессов при различии внешних влияний; М. Биша - как совокупность функций, сопротивляющихся смерти; И.П. Павлов — как сложный химический процесс; Ф. Энгельс - как способ существования белковых тел, значимым моментом которого является обмен веществ с окружающей средой. Одно из наиболее полных определений жизни с учетом современного уровня знаний дал отечественный ученый М.В. Волькенштейн. По его мнению, существующие на Земле живые тела представляют собой открытые саморегулирующиеся и самовоспроизводящиеся системы, построенные из биополимеров - белков и нуклеиновых кислот. Здесь подчеркивается значение нуклеиновых кислот, обеспечивающих преемственность признаков и свойств.
Живым организмам присущи определенные свойства. Часто эти свойства в той или иной степени характерны и для неживой природы, что подчеркивает единство эволюционных процессов. Однако проявление этих свойств и их совокупность не схожи у живых и неживых объектов. Именно это - совокупность и характер проявления — как раз и определяет сущность жизни. Рассмотрим ряд свойств живых организмов в сравнении со свойствами неживых объектов [19, 20].
◊ Единство химического состава. В состав живых организмов входят те же химические элементы, что и в объекты неживой природы. Однако соотношение элементов в живом и неживом неодинаково. Элементный состав неживой природы наряду с кислородом представлен в основном кремнием, железом, магнием, алюминием и т.д. В живых организмах 98% химического состава приходится на четыре элемента: углерод, кислород, азот и водород, и, кроме того, живые организмы построены в основном из четырех крупных групп сложных органических молекул — биологических полимеров: нуклеиновых кислот, белков, полисахаридов, жиров, которые редко встречаются в неживой природе.
◊ Обмен веществ. Все живые организмы способны к обмену веществ с окружающей средой - поглощают из нее необходимые вещества и выделяют продукты жизнедеятельности. Обмен веществ - двусторонний процесс: во-первых, в результате ряда сложных химических превращений вещества из окружающей среды уподобляются органическим веществам живого организма и из них строится его тело; во-вторых, сложные органические соединения распадаются на простые, при этом утрачивается их сходство с веществами организма и выделяется энергия, необходимая для реакций биосинтеза. Обмен веществ обеспечивает постоянство химического состава и строения всех частей организма и, как следствие, постоянство их функционирования в непрерывно меняющихся условиях окружающей среды, т.е. обеспечивает гомеостаз. В неживой природе также существует обмен веществ. Но небиологический круговорот веществ сводится к простому переносу их с одного места на другое или изменению их агрегатного состояния, например превращению воды в пар или лед.
◊ Самовоспроизведение (репродукция) и наследственность. При размножении живых организмов потомство обычно похоже на родителей: кошки воспроизводят котят, собаки - щенят, из семян одуванчика вырастает одуванчик. Таким образом, размножение - это свойство организмов воспроизводить себе подобных. В основе самовоспроизведения лежит образование новых молекул и структур на основе информации, заложенной в ДНК. Благодаря репродукции не только целые организмы, но и клетки, органоиды клеток после деления сходны со своими предшественниками; так, из одной молекулы ДНК при ее удвоении образуются две дочерние молекулы, полностью повторяющие исходную. Следовательно, самовоспроизведение тесно связано с наследственностью - способностью организмов обеспечивать передачу признаков, свойств, особенностей развития из поколения в поколение, что обусловливает преемственность поколений.
◊ Изменчивость, развитие и рост. Под изменчивостью понимают способность организмов приобретать новые признаки и свойства на основе изменения молекул ДНК. Изменчивость создает разнообразный материал для естественного отбора и соответственно предпосылки для развития и роста живых организмов. Развитие - необратимое направленное закономерное изменение объектов живой и неживой природы. В результате развития возникает новое качественное состояние систем. Развитие живой формы существования материи представлено индивидуальным развитием организмов (онтогенез) и историческим развитием видов (филогенез). В процессе развития постепенно и последовательно формируется специфическая структурная организация живого организма, а увеличение его массы обусловлено репродукцией макромолекул, элементарных структур клеток и самих клеток. Филогенез, или эволюция в целом, - это необратимое и направленное развитие живой природы, сопровождающееся образованием новых видов и прогрессивным усложнением жизни. Результатом эволюции является все многообразие живых организмов на Земле.
◊ Раздражимость и энергозависимость. Любой организм неразрывно связан с окружающей средой: извлекает из нее необходимые вещества, подвергается воздействию неблагоприятных факторов среды, вступает во взаимодействие с другими организмами и т.д. В процессе эволюции у живых организмов выработалось и закрепилось свойство раздражимости — избирательной реакции на внешние воздействия. Всякое изменение окружающих организм условий среды представляет собой по отношению к нему раздражение, а реакция организма на внешние раздражители служит показателем его чувствительности и проявлением раздражимости. Кроме того, живые организмы обладают свойством энергозависимости; это открытые для поступления энергии системы, устойчивые лишь при условии непрерывного доступа к ним энергии и материи извне. Живые организмы существуют до тех пор, пока получают энергию и материю из окружающей среды.
◊ Ритмичность - еще одно следствие тесного взаимодействия живой и неживой природы. В природе повсюду распространены колебательные процессы: океанские приливы и отливы, смена дня и ночи, фаз луны, чередование времен года, периодическое увеличение солнечной активности, цикличность геологических процессов. Периодические изменения в окружающей среде оказывают существенное влияние на живую природу и на собственные ритмы живых организмов. В живых системах ритмичность проявляется в периодических изменениях интенсивности физиологических функций с различными периодами колебаний (от нескольких секунд до года и столетия): суточные ритмы сна и бодрствования у человека, сезонные ритмы активности и спячки у некоторых млекопитающих (суслики, ежи, медведи) и др. Ритмичность обеспечивает согласование функций организма и окружающей среды, т.е. приспособление к периодически изменяющимся условиям существования. Например, сезонные и суточные ритмы выработались как приспособление живых организмов к геофизическим циклам среды.
◊ Саморегуляция (авторегуляция) - способность живых организмов, обитающих в непрерывно меняющихся условиях окружающей среды, поддерживать постоянство своего химического состава и интенсивность физиологических процессов. При этом недостаток поступления каких-либо питательных веществ мобилизует внутренние ресурсы организма, а при избытке какого-либо вещества его синтез прекращается. Например, понижение концентрации такого фермента, как АТФ - универсального аккумулятора энергии в клетке, служит сигналом, запускающим процесс его синтеза; при восполнении запаса АТФ синтез его прекращается. Уменьшение количества клеток в ткани (например, в результате травм) вызывает усиленное размножение оставшихся клеток; восстановление количества клеток до нормального дает сигнал о прекращении интенсивного клеточного деления.
◊ Дискретность (прерывистость, разделенностъ). Жизнь на Земле проявляется в виде дискретных форм, т.е. отдельный организм или биологическая система (вид, биогеоценоз и др.) состоит из обособленных или отграниченных в пространстве, но тем не менее тесно связанных и взаимодействующих между собой частей, образующих структурно-функциональное единство. Например, любой вид включает отдельные особи; тело высокоорганизованной особи образуют пространственно отграниченные органы, которые в свою очередь состоят из отдельных тканей и клеток; энергетический аппарат клетки представлен отдельными митохондриями, аппарат синтеза белка - рибосомами2 и далее вплоть до макромолекул, каждая из которых может выполнять функцию, лишь будучи пространственно изолированной от других. Дискретность строения организма - основа его структурной упорядоченности. Она создает возможность постоянного самообновления его путем замены «износившихся» структурных элементов (молекул, ферментов, органоидов клетки3, целых клеток) без прекращения выполняемой функции. Дискретность вида предопределяет возможность его эволюции через гибель или устранение от размножения неприспособленных особей и сохранения особей с полезными для выживания признаками.
Уровни организации жизни
Мир живой природы представляет собой совокупность биологических систем разного уровня организации и различной соподчиненности. Обычно выделяют несколько уровней организации живой материи [6, 20, 28].
◊ Молекулярный уровень. Различие живых и неживых систем проявляется уже на молекулярном уровне. На уровне функционирования биологических макромолекул (нуклеиновых кислот, белков, полисахаридов), а также других важных органических веществ начинаются важнейшие процессы жизнедеятельности организма - обмен веществ и превращение энергии, передача наследственной информации и др. В настоящее время выясняется, каким образом и в какой мере рост и развитие организмов, хранение и передача наследственной информации, превращение энергии в живых клетках и другие явления обусловлены структурой и свойствами биологически важных макромолекул (главным образом белков и нуклеиновых кислот). Безусловный интерес представляет так называемая репликация - удвоение молекул ДНК (у некоторых вирусов РНК) при участии специальных ферментов, которая обеспечивает точное копирование генетической информации, заключенной в молекулах ДНК, и передачу ее от поколения к поколению.
◊ Клеточный уровень. Клетка — структурная и функциональная единица, а также единица размножения и развития всех живых организмов, обитающих на Земле. Она может существовать как отдельный организм (бактерии, простейшие, некоторые водоросли и грибы), так и в составе тканей многоклеточных животных, растений, грибов. Единственное исключение из этого правила — вирусы, представляющие собой неклеточные формы жизни, но вирусы могут проявлять свойства живых систем только в клетках. Содержимое клетки именуется протоплазмой. В каждой клетке имеется генетический аппарат, который обычно заключен в ядре, отделенном мембранами от цитоплазмы.
Клетки различны по величине; так, клетки некоторых бактерий имеют диаметр 0,1-0,23 мкм, а диаметр яйца страуса в скорлупе достигает 155 мм; диаметр большинства эукариотных клеток (с оформленным клеточным ядром, т.е. все клетки, кроме бактерий) составляет 10-100 мкм. Многообразные функции клеток выполняются специализированными внутриклеточными структурами - органоидами. Универсальные органоиды эукариотных клеток в ядре - хромосомы, в цитоплазме - рибосомы, митохондрии, эндоплазматическая сеть, комплекс Гольджи, лизосомы, клеточная мембрана. Во многих клетках присутствуют мембранные структуры, способствующие поддержанию формы клетки.
Важнейшими химическими компонентами клетки считаются белки, включая ферменты, которые содержатся как в самой клетке, так и в жидких средах организма. Однако синтезируются они только в клетке, которая осуществляет пространственную организацию химических процессов. Так, процесс клеточного дыхания у всех клеток, кроме бактерий, происходит только на мембранах митохондрий, а синтез белка — на рибосомах. Благодаря концентрации ферментов, упорядоченному их расположению в структурах ускоряются реакции, организуется их сопряжение (принцип конвейера), разделяются разнородные процессы. Для строения клетки характерна микрогетерогенность (микронеоднородность), которая позволяет синтезировать различные вещества из одних и тех же предшественников в одно время в общем микрообъеме. Принцип компактности, присущий метаболизму клетки, особенно выражен в структуре ДНК. Например, ДНК яйцеклетки человека весит 6 · 10~12 г и при этом кодирует свойства всех белков человека. Внутри клетки непрерывно поддерживается определенная концентрация ионов, отличная от их концентрации в окружающей среде, в результате чего образуются выпячивания клеточной мембраны, которые могут замыкаться и отделяться внутрь клетки в виде пузырьков. Клетки способны захватывать из окружающей их среды капельки с крупными молекулами, включая белки или даже вирусы и небольшие клетки.
У всех клеток одного организма геном (совокупность генов) не отличается по объему потенциальной информации от генома оплодотворенной яйцеклетки. Это доказывают опыты с пересадкой ядра узкоспециализированной клетки в цитоплазму энуклеированной яйцеклетки, после чего может развиться нормальный организм. Различия свойств клетки многоклеточного организма объясняются неодинаковой активностью генов, что обусловливает дифференцировку клеток. В результате одни клетки становятся возбудимыми (нервные), другие приобретают сократимые белки, образующие миофибриллы (мышечные), третьи синтезируют пищеварительные ферменты или гормоны (железистые) и т.д. Многие клетки полифункциональны; так, клетки печени синтезируют различные белки плазмы крови и желчь, накапливают гликоген и превращают его в глюкозу, окисляют чужеродные вещества (в том числе и многие лекарства). Во всех клетках активны гены общеклеточных функций. Следовательно, сходных признаков в разных клетках значительно больше, чем специальных признаков. Клетки близкого происхождения и сходных функций образуют ткани.
В организме человека около 1014 клеток. В некоторых тканях количество клеток остается постоянным в течение всей жизни организма. В этих тканях делятся относительно малодифференцированные клетки, резерв которых самоподдерживается, а одна из дочерних клеток дифференцируется. Например, у человека ежедневно погибает около 70 млрд клеток кишечного эпителия и 2 млрд эритроцитов. Во многих других тканях в клеточный цикл входят вполне дифференцированные клетки, и тогда деление клетки не может завершиться до конца, а ограничивается удвоением хромосом или вообще не начинается, и клетка выходит из цикла после некоторого момента. Некоторые ядра не входят в цикл в течение всей жизни дифференцированной клетки (нейроны, волокна скелетных мышц), при этом продолжительность жизни клетки соответствует жизни организма. Минимальная продолжительность жизни клетки (кишечного эпителия) человека -1-2 дня.
Во всех клетках происходит интенсивное обновление веществ и структур. Огромное количество клеток в каждой ткани, объединенных метаболическими и регуляторными процессами, их постоянное внутреннее обновление обеспечивают надежность работы органов многоклеточного организма.
На клеточном уровне основное внимание ученые уделяют проблемам морфологической организации клетки, специализации клеток в ходе развития, функциям клеточной мембраны механизмов и регуляции деления клетки, строению и функциям таких органоидов, как хромосомы, митохондрии, рибосомы, а также другим включениям клетки.
◊ Организменный уровень. Многоклеточный организм представляет собой целостную систему органов и тканей, специализированных для выполнения различных функций. Ткань - совокупность сходных по строению клеток, объединенных выполнением общей функции. Орган - это структурно-функциональное объединение нескольких типов тканей. Например, кожа человека как орган включает эпителий и соединительную ткань, которые вместе выполняют целый ряд функций; из них наиболее значительная — защитная. В рамках организменного (онтогенетического) уровня изучают особь и свойственные ей как целому черты строения и физиологические процессы — механизмы адаптации (акклиматизации) и поведения (в том числе функции центральной нервной системы), а также такие актуальные проблемы, как дифференцировка тканей, т.е. превращение в процессе индивидуального развития организма (онтогенеза) первоначально одинаковых, неспециализированных клеток зародыша в специализированные клетки тканей и органов,
О Популяционно-видовой уровень. Популяция как система надорганизменного порядка - это совокупность организмов одного и того же вида, объединенная общим местом обитания. В данной системе осуществляются простейшие, элементарные эволюционные преобразования. Популяционная биология изучает факторы, влияющие на численность популяции, проблемы сохранения исчезающих видов, динамики генетического состава популяций, действие факторов микроэволюции и т.д. Для хозяйственной деятельности человека важны такие проблемы популяционной биологии, как контроль численности видов, поддержание оптимальной численности эксплуатируемых и охраняемых популяций.
◊ Биогеоценотически-биосферный уровень. Биогеоценоз - совокупность организмов разных видов и различной сложности организации со всеми факторами среды обитания.
Биосфера - самая крупная единица организации живой материи на Земле. Соответствующая отрасль биологии изучает проблемы взаимоотношений организмов в биоценозах, условия и факторы, определяющие их численность, продуктивность и устойчивость, влияние деятельности человека на сохранение биоценозов и их комплексов. На втором наиболее глобальном (биосферном) подуровне биология решает такие глобальные проблемы, как определение интенсивности образования свободного кислорода растительным покровом Земли, изменения концентрации углекислого газа в атмосфере, связанного с деятельностью человека.
§ 9.2. Концепции возникновения жизни
Подходы к решению проблемы возникновения жизни
Вопрос о возникновении жизни беспокоит человечество достаточно давно. В число теорий, получивших весьма широкое распространение и доминировавших в те или иные периоды развития естествознания, обычно включают: теорию стационарного состояния, в соответствии с которой жизнь существовала всегда; креационизм, утверждающий, что жизнь создана сверхъестественным существом в результате акта творения; теорию самопроизвольного зарождения, т.е. жизнь возникала и возникает неоднократно из неживого вещества; теорию панспермии, утверждающую, что жизнь занесена на Землю извне; теорию биохимической эволюции. Следует отметить, что теории стационарного состояния и креационизм возникли под воздействием преимущественно религиозных представлений о мироздании; креационизм в настоящее время (в той или иной форме) поддерживается сторонниками божественного сотворения мира.
◊ Теорию самозарождения, или самопроизвольного возникновения жизни, обычно связывают с развитием собственно естественно-научных представлений о возникновении жизни на Земле. Сторонники этой теории находили подтверждение своей идеи в опытах с появлением червей в гниющем мясе, земле, отбросах, мышей в мешках с зерном и т.д. Данную точку зрения разделяли Аристотель, Г. Галилей, И.В. Гёте, Р. Декарт. Их авторитет долгое время поддерживал теорию самозарождения на главенствующих позициях в естествознании.
Одним из первых эту точку зрения подверг сомнению итальянский врач и естествоиспытатель Ф. Реди (XVII в.), показав, что в хорошо изолированном от окружающей среды мясе черви не появляются. Он же провозгласил принцип «все живое от живого». Однако окончательно этот принцип получил распространение лишь в 1860-е гг., что связано с опытами Л. Пастера, который доказал его на микробиологическом уровне.
◊ Теория панспермии объясняла зарождение жизни на Земле естественными причинами. Видоизменяясь, эта теория существовала с древности до начала XX в. По представлениям ее сторонников, таких известных ученых, как С.А. Аррениус, Г. Гельмгольц, В.И. Вернадский, зародыши жизни (например, споры микроорганизмов) рассеяны в мировом пространстве и переносятся с одного небесного тела на другое с метеоритами или под действием давления света; следовательно, жизнь занесена на Землю из Космоса. Эти представления снимали противоречия, характерные для гипотезы самозарождения. Однако оставались открытыми вопросы о том, где и как зародилась жизнь и каким образом она была перенесена на Землю. Особенно остро эти вопросы встали после открытия космических лучей и выяснения действия радиации на биологические объекты. В настоящее время периодически предпринимаются попытки возродить эту теорию.
В начале XX в. накопившиеся противоречия, с которыми не могли справиться теории самозарождения и панспермии, привели к дальнейшим поискам объяснения способа возникновения жизни на Земле. Так, Г. Миллер выдвинул гипотезу о случайном возникновении первичной молекулы живого вещества. Однако было подсчитано, что это событие может произойти с вероятностью, равной вероятности того, что обезьяна отпечатает сонет У. Шекспира на пишущей машинке, если будет продолжительное время ударять по клавишам.
◊ Теория биохимической эволюции отражает современные представления о происхождении жизни на Земле и основывается на работах А.И. Опарина (1924) и Дж. Холдейна (1929). Эти представления заключаются в признании абиогенного, т.е. небиологического, возникновения органических веществ из химических элементов и неорганических молекул, составлявших атмосферу Земли в древности, путем длительной молекулярной эволюции. За прошедшие десятилетия концепция биохимической эволюции была подтверждена и существенно дополнена новыми данными из разных областей естествознания.
Условия и факторы зарождения жизни
В настоящее время известно, что все живые существа, во - пеpвых, обладают совокупностью одних и тех же свойств и состоят из одних и тех же групп биологических полимеров, выполняющих определенные функции; во-вторых, последовательность биохимических превращений, обеспечивающих обменные процессы, у них сходна вплоть до деталей. Например, расщепление глюкозы, биосинтез белка и другие реакции у самых разных организмов протекают почти одинаково. Следовательно, вопрос о происхождении жизни сводится к тому, как и в каких условиях возникла столь универсальная система биохимических превращений [13, 20, 23, 29].
Несмотря на общность происхождения планет Солнечной системы, только на Земле появилась жизнь и достигла исключительного многообразия. Связано это с тем, что для возникновения жизни необходимы некоторые космические и планетарные условия. Во-первых, масса планеты не должна быть слишком большой, так как энергия атомного распада природных радиоактивных веществ может привести к перегреванию планеты или радиоактивному загрязнению среды, не совместимому с жизнью; слишком маленькие планеты не могут удержать около себя атмосферу, потому что сила притяжения их невелика. Во-вторых, планета должна вращаться вокруг звезды по круговой или близкой к круговой орбите, что позволит постоянно и равномерно получать от нее необходимое количество энергии. В-третьих, интенсивность излучения светила должна быть постоянной; неравномерность потока энергии будет препятствовать возникновению и развитию жизни, поскольку существование живых организмов возможно в узких температурных пределах. Всем этим условиям удовлетворяет Земля, на которой около 4,6 млрд лет назад начали создаваться условия для возникновения жизни.
На первых этапах формирования Земли тяжелые элементы перемещались к ее центру, а более легкие оставались на поверхности. Металлы и другие способные окисляться элементы соединялись с кислородом, и в атмосфере Земли не было свободного кислорода. Атмосфера состояла из свободного водорода и его соединений (Н2О, СН4, NH3, HCN), т.е. носила восстановительный характер. По мнению А.И. Опарина, это служило важной предпосылкой для возникновения органических молекул небиологическим путем. До середины XX в. многие ученые полагали, что эти соединения могут возникать только в живом организме. Именно поэтому их назвали органическими соединениями в противоположность веществам неживой природы - минералам, названным неорганическими соединениями. Однако в 1953 г. Л.С. Миллер экспериментально доказал возможность абиогенного синтеза органических соединений из неорганических. Пропуская электрический разряд через смесь Н2, Н2О, СН4 и NH3, он получил набор нескольких аминокислот и органические кислоты. Позднее было установлено, что аналогичным путем в отсутствие кислорода могут быть синтезированы очень многие органические соединения, входящие в состав биологических полимеров (белков, нуклеиновых кислот и полисахаридов).
Возможность абиогенного синтеза органических соединений подтверждается тем, что в космическом пространстве обнаружены цианистый водород, формальдегид, муравьиная кислота, метиловый и этиловый спирты и др. В некоторых метеоритах обнаружены жирные кислоты, сахара, аминокислоты. Все это свидетельствует о том, что достаточно сложные органические соединения могли возникать в условиях, существовавших на Земле 4,0-4,5 млрд лет назад.
Более 4 млрд лет назад извергалось множество вулканов с выбросом огромного количества раскаленной лавы, выделялись большие объемы пара, сверкали молнии. По мере остывания планеты водяные пары, находившиеся в атмосфере, конденсировались и обрушивались на Землю ливнями, образуя огромные водные пространства. Поскольку поверхность Земли в то время была горячей, вода испарялась, а затем, охлаждаясь в верхних слоях атмосферы, вновь выпадала на поверхность планеты. Это продолжалось в течение многих миллионов лет. В водах первичного океана были растворены компоненты атмосферы, различные соли. Кроме того, туда попадали и непрерывно образующиеся в атмосфере под действием жесткого ультрафиолетового излучения Солнца, высокой температуры в областях грозовых разрядов и активной вулканической деятельности органические соединения - сахара, аминокислоты, азотистые основания, органические кислоты и др.
По мере смягчения условий на Земле стало возможным образование сложных органических соединений благодаря, вероятно, поглощению различных веществ на поверхности глин и других неорганических осадков, что повышало концентрацию реагирующих между собой веществ, в результате чего появились первичные биополимеры - полипептиды и полинуклеотиды.
Таким образом, условиями для абиогенного возникновения органических соединений были: восстановительный характер атмосферы Земли (соединения, обладающие восстановительными свойствами, легко вступают во взаимодействия между собой и веществами-окислителями), высокая температура, грозовые разряды и мощное ультрафиолетовое излучение Солнца, которое тогда еще не задерживалось озоновым экраном.
Первичный океан, по-видимому, содержал в растворенном виде различные органические и неорганические молекулы, попавшие в него из атмосферы и вымывавшиеся из поверхностных слоев Земли. Концентрация органических соединений постоянно увеличивалась, и в конце концов воды океана стали «бульоном» из белковоподобных веществ - пептидов, а также нуклеиновых кислот и других органических соединений.
Органические молекулы имеют большую молекулярную массу и сложную пространственную конфигурацию. Они окружены водной оболочкой и объединяются, образуя высокомолекулярные комплексы -- коацерваты, или коацерватные капли (как их называл А.И. Опарин). Коацерваты обладали способностью поглощать различные вещества, растворенные в водах первичного океана. В результате этого внутреннее строение коацервата изменялось, что вело или к его распаду, или к накоплению веществ, т.е. к росту и изменению химического состава, повышающему устойчивость коацерватной капли в постоянно меняющихся условиях.
В массе коацерватных капель происходил отбор наиболее устойчивых в данных конкретных условиях. Достигнув определенных размеров, материнская коацерватная капля могла распадаться на дочерние, но продолжали существовать только те дочерние коацервагаые капли, которые, вступая в элементарные формы обмена со средой, сохраняли относительное постоянство своего состава. В дальнейшем они приобрели способность поглощать из окружающей среды не всякие вещества, а лишь те, которые обеспечивали их устойчивость, а также выделять наружу продукты обмена. Параллельно увеличивались различия между химическим составом капли и окружающей среды. В процессе длительного отбора (химической эволюции) сохранились лишь те коацерваты, которые при распаде на дочерние не утрачивали особенностей структуры, т.е. приобретали свойство самовоспроизведения. В ходе эволюции у важнейших составных частей коацерватных капель полипептидов — выработалась способность к каталитической активности, т.е. к значительному ускорению биохимических реакций, приводящих к превращению органических соединений, а полинуклеотиды оказались способными связываться друг с другом по принципу дополнения (комплиментарности) и, следовательно, осуществлять неферментативный синтез дочерних полинуклеотидных цепей.
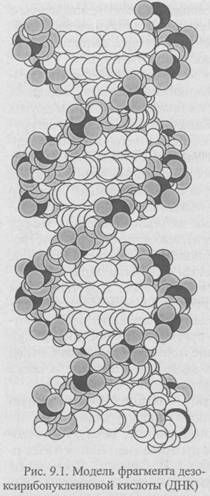
Следующий важный шаг предбиологической эволюции — объединение способности полинуклеотидов к самовоспроизведению с возможностью полипептидов ускорять течение химических реакций, так как удвоение молекул ДНК эффективнее осуществляется при участии белков, обладающих каталитической активностью. Однако стабильность «удачных» комбинаций аминокислот в полипептидах может обеспечиваться только при сохранении информации о них в нуклеиновых кислотах. Связь нуклеиновых кислот и белковых молекул в конце концов привела к возникновению генетического кода, т.е. такой организации молекул ДНК (рис. 9.1), в которой последовательность нуклеотидов стала служить информацией для построения конкретной последовательности аминокислот в белках.
Дальнейшая прогрессивная эволюция предбиологических структур была возможна только при усложнении обменных процессов и пространственном разделении различных синтетических и энергетических процессов внутри коацервата. Более прочную изоляцию внутренней среды от внешних воздействий по сравнению с той, которую обеспечивала водная оболочка, могла осуществить лишь биологическая мембрана. Вокруг коацерватов, богатых органическими соединениями, возникли слои липидов, отделивших коацерват от окружающей водной среды. В процессе эволюции липиды преобразовались в наружную мембрану, значительно повысившую жизнеспособность и устойчивость организмов. Появление мембраны, отделяющей содержимое коацервата от окружающей среды и обладающей способностью к избирательной проницаемости, предопределило направление дальнейшей химической эволюции по пути развития все более совершенных саморегулирующихся систем вплоть до возникновения первых клеток.
Начальные этапы биологической эволюции
Образование первичных клеточных организмов положило начало биологической эволюции. Считается, что отбор коацерватов и пограничный этап химической и биологической эволюции продолжались около 750 млн лет. В конце этого периода появились первые примитивные безъядерные клетки — прокариоты. Первые живые организмы — гетеротрофы - использовали в качестве источника энергии (пищи) органические соединения, растворенные в водах первичного океана. Поскольку в атмосфере Земли не было свободного кислорода, гетеротрофы имели анаэробный (бескислородный) тип обмена веществ, эффективность которого невысока. Увеличение количества гетеротрофов привело к истощению вод первичного океана, где оставалось все меньше готовых органических веществ, которые можно было использовать для питания.
В более выгодном положении оказались организмы, которые развили способность использовать энергию солнечного излучения для синтеза органических веществ из неорганических — фотосинтеза. Таким образом, появился принципиально новый источник питания. Например, современные фотосинтезирующие пурпурные бактерии благодаря солнечному излучению окисляют сероводород до сульфатов. Высвобождающийся в результате реакции окисления водород идет на восстановление диоксида углерода до углеводов с образованием воды. Использование органических соединений в качестве источника (донора) водорода привело к появлению автотрофных организмов (способных синтезировать из неорганических веществ все необходимые для жизни органические вещества).
Следующий шаг эволюции связан с развитием у фотосинтезирующих организмов способности использовать воду в качестве источника водорода для синтеза органических молекул. Усвоение углекислого газа такими организмами сопровождалось выделением кислорода и включением углерода в органические соединения. Так в атмосфере Земли начал накапливаться кислород. Первыми фотосинтезирующими организмами, выделяющими в атмосферу кислород, были цианобактерии (цианеи).
Переход от первичной атмосферы к среде, содержащей кислород, представляет собой важнейшее событие как в эволюции живых существ, так и в преобразовании минералов. Во - пеpвых, кислород, выделяющийся в атмосферу, в верхних ее слоях под действием мощного ультрафиолетового излучения Солнца превращается в активный озон (О3), который способен поглощать большую часть жестких коротковолновых ультрафиолетовых лучей, разрушительно действующих на сложные органические соединения. Во-вторых, в присутствии свободного кислорода возможен кислородный тип обмена веществ, энергетически более выгодный. Образование свободного кислорода вызвало к жизни многочисленные новые формы аэробных живых организмов и более широкое использование ими ресурсов окружающей среды.
В результате взаимополезного симбиоза различных прокариотических (не обладающих оформленным клеточным ядром) клеток возникли ядерные, или эукариотические, организмы (эукариоты). Основой симбиоза была, вероятно, гетеротрофная амебоподобная клетка. Питанием для нее служили более мелкие клетки и, в частности, дышащие кислородом аэробные бактерии, способные функционировать и внутри клетки-хозяина, производя энергию. Те крупные амебовидные клетки, в теле которых аэробные бактерии оставались невредимыми, оказались в более выгодном положении, чем клетки, получавшие энергию анаэробным путем — брожением. В дальнейшем бактерии-симбионты превратились в митохондрии (органеллы клеток, где протекают реакции, обеспечивающие клетки энергией). Когда к поверхности клетки-хозяина прикрепилась вторая группа симбионтов - жгутикоподобных бактерий, сходных с современными спирохетами, подвижность и способность к нахождению пищи такого организма резко возросли. Так возникли примитивные животные клетки - предшественники нынешних жгутиковых простейших.
Образовавшиеся подвижные эукариоты путем симбиоза с фотосинтезирующими (возможно, цианобактериями) организмами дали водоросль, или растение, причем строение пигментного комплекса у фотосинтезирующих анаэробных бактерий сходно с пигментами зеленых растений. Такое сходство указывает на возможность эволюционного преобразования фотосинтезирующего аппарата анаэробных бактерий в аналогичный аппарат зеленых растений.
Изложенную гипотезу о возникновении эукариотических клеток через ряд последовательных симбиозов приняли многие современные ученые, поскольку она хорошо обоснованна. Во-первых, одноклеточные водоросли и сейчас легко вступают в союз с животными - эукариотами; например, в теле инфузории туфельки обитает водоросль хлорелла. Во-вторых, некоторые органоиды клетки — митохондрии и пластиды — по строению ДНК очень похожи на прокариотические клетки-бактерии и цианобактерии.
Возможности эукариот по использованию среды существенно выше, чем у прокариот, поскольку они имеют диплоидный (двойной) набор генов. У прокариот любая мутация сразу проявляется в виде признака. Если мутация полезна, организм продолжает существовать, если вредна - он погибает, т.е. прокариоты непрерывно приспосабливаются к изменениям окружающей среды, но лишены возможности формировать крупные структурные изменения. Появление двойного набора генов у эукариот сделало возможным накопление непроявляющихся фенотипических мутаций и, следовательно, формирование резерва наследственной изменчивости - основы эволюционных преобразований.
Возможности одноклеточных в освоении среды обитания были ограничены, так как дыхание и питание простейших осуществляются через поверхность тела. При увеличении размеров клетки одноклеточного организма его поверхность возрастает по квадратичному закону, а объем - по кубическому, поэтому биологическая мембрана, окружающая клетку, не могла обеспечивать кислородом слишком большой организм. Иной эволюционный путь осуществился позже, около 2,6 млрд лет назад, когда появились многоклеточные организмы, эволюционные возможности которых значительно шире.
Первая гипотеза о происхождении многоклеточных организмов принадлежит Э. Геккелю (вторая половина XIX в.). При ее построении он исходил из исследований эмбрионального развития ланцетника (род животных класса бесчерепных), проведенных А.О. Ковалевским и другими зоологами. Геккель полагал, что начальная стадия развития зародыша (стадия зиготы) соответствует одноклеточным предкам, а стадия развития зародыша многоклеточных животных в процессе бластуляции (заключительной фазе периода дробления яйца) — шарообразной колонии жгутиковых. В дальнейшем, согласно этой гипотезе, произошло впячивание (инвагинация) одной из сторон шарообразной колонии и образовался гипотетический двухслойный организм, названный Геккелем гастреей. Теория Геккеля сыграла важную роль в истории науки, способствуя утверждению монофилетических (т.е. из одного корня) представлений о происхождении многоклеточных.
Основу современных представлений о возникновении многоклеточных организмов составляет гипотеза фагоцителлы И.И. Мечникова. По его представлениям, многоклеточные произошли от колониальных простейших - жгутиковых. Пример такой организации - ныне существующие колониальные жгутиковые типа вольвокс. Среди клеток колонии выделяются движущиеся, снабженные жгутиками, фагоцитирующие добычу и уносящие ее внутрь колонии, и половые, функцией которых является размножение. Так колония превратилась в примитивный, но целостный многоклеточный организм. О справедливости гипотезы фагоцителлы говорит строение примитивного многоклеточного организма - трихоплакса, который по строению соответствует гипотетической фагоцителле и поэтому должен быть выделен в особый тип животных - фагоцителлоподобных, заполняющих брешь между многоклеточными и одноклеточными организмами [18].
Таким образом, в настоящее время большинство исследователей в области естествознания признает, что возникновение жизни на Земле связано с длительным процессом химической эволюции. Формирование структуры, отграничивающей организм от окружающей среды, — мембраны с присущими ей свойствами способствовало появлению живых организмов и ознаменовало начало биологической эволюции. Как простейшие живые организмы, возникшие около 3 млрд лет назад, так и устроенные более сложно в основе своей структурной организации имеют клетку.
§ 9.3. Эволюционное учение в биологии
Становление эволюционного учения
Развитие эволюционных идей в биологии имеет достаточно длительную историю [И, 12, 19, 20, 26, 33, 38]. Так, уже в античности Анаксимен (VI в. до н.э.) говорил о внезапном превращении видов, а Эмпедокл (V в. до н.э.) считал, что сложные организмы образуются на основе случайного сочетания отдельных органов, причем нежизнеспособные сочетания вымирают, а удачные сохраняются. В средние века представления о живой природе базировались на теории постепенного развития живых форм (от простого к сложному), созданной Аристотелем (IV в. до н.э.).
Начиная с XIV в. число ученых, принимавших идеи эволюции органического мира, постоянно возрастало. В XVIII в. развитию этих идей способствовало создание систематики растительного и животного мира К. Линнеем (хотя сам автор этой систематики выступал в защиту постоянства видов и креационизма), включившем в нее всех известных тогда животных и все известные растения. Он описал более 8000 видов растений, установил единообразную терминологию и порядок описания видов. В основу своей классификации он положил принцип иерархичности, т.е. соподчиненности, таксонов (систематических единиц). В системе Линнея самым крупным таксоном был класс, самым мелким - вид.
В конце XVIII - начале XIX в. стали складываться собственно эволюционные концепции, характеризующиеся использованием исторического подхода к объяснению целесообразности живого. Считается, что первой целостной эволюционной концепцией является ламаркизм, тесно связанный с развитием трансформизма в истории эволюционного учения. Ж.Б. Ламарк высказал идею о том, что организмы изменчивы, а виды (и другие таксономические категории) условны и постепенно преобразуются в новые виды. Общей тенденцией исторических изменений организмов он считал постепенное совершенствование их организации, движущей силой которой является изначальное (заложенное Творцом) стремление природы к прогрессу. Более того, организмам присуща изначальная способность целесообразно реагировать на изменения внешних условий, а изменения организмов, приобретенные в течение жизни в ответ на изменения условий, наследуются. Таким образом, в своей теории Ламарк впервые объединил идею изменяемости видов и идею прогрессивной эволюции.
Ж. Кювье разработал теорию катастроф, используя палеонтологическую летопись Земли. Он истолковывал ископаемые останки как результаты «катастроф» в ее истории, порождающих новые виды. Важно отметить, что в геологическом прошлом Земли действительно были определенные отрезки времени, когда сравнительно быстро (по геологическим меркам) исчезали одни виды и возникали другие. Это заставляет периодически возвращаться к идеям Кювье на другом уровне познания в рамках теории неокатастрофизма.
Величайшим событием в изучении феномена жизни и ее эволюции стала теория Ч. Дарвина. В своей работе «Происхождение видов путем естественного отбора» (1859) он, обобщив отдельные эволюционные идеи, создал стройную теорию эволюции. Движущими силами эволюции он считал наследственную изменчивость и естественный отбор, а элементарной единицей эволюции - вид. Дарвин сформулировал следующие положения. Во-первых, изменчивость свойственна любой группе животных и растений и организмы различаются во многих отношениях. Во-вторых, число организмов каждого вида, рождающихся на свет, больше того их числа, которое может найти пропитание и выжить, однако численность каждого вида в естественных условиях довольно постоянна. В-третьих, поскольку рождается больше особей, чем может выжить, происходит борьба за существование, конкуренция за пищу и местообитание. В-четвертых, изменения организма, облегчающие его выживание в определенной среде, дают своему обладателю преимущество перед другими организмами, менее приспособленными к внешним условиям; идея выживаемости наиболее приспособленных организмов является ядром теории естественного отбора. В-пятых, выживающие особи дают начало следующему поколению, благодаря чему «удачные» положительные изменения передаются последующим поколениям.
Параллельно с теорией Дарвина развивались концепции, которые частично или полностью строились на других идеях. Так, в конце XIX — начале XX в. сложилась теория неоламаркизма, представленная тремя основными течениями: ортоламаркизм, механоламаркизм и психоламаркизм. Представители ортоламаркизма полагали, что направленность эволюции обусловлена внутренними изначальными свойствами организмов. Эти взгляды родственны идеям автогенезиса, который рассматривает эволюцию как процесс развертывания предсуществующих задатков, носящий целенаправленный характер и происходящий на основе изначальных внутренних потенциальных возможностей.
Механоламаркизм объясняет эволюционные преобразования организмов их изначальной способностью целесообразно реагировать на изменения внешней среды (эктогенез), изменяя структуры и функции. Эта концепция основана на признании так называемой адекватной соматической индукции, т.е. адаптивные модификации являются эволюционными новообразованиями и наследуются. Следует заметить, что эктогенез противопоставлялся автогенезу.
Психоламаркизм рассматривает в качестве причин эволюции сознательные волевые акты организмов, которые присущи не только животным, но и составляющим их клеткам. Поэтому эволюция трактуется как постепенное усиление роли сознания в развитии от примитивных существ до разумных форм жизни. Данная концепция способствовала развитию учения о панпсихизме (всеобщей одушевленности).
Из других концепций конца XIX - начала XX в., которые не опирались на теорию Дарвина, следует упомянуть телеогенез, сальтационизм, генетический антидарвинизм. Телеогенез (телеологическая концепция эволюции) в сущности близок ортоламаркизму, так как исходит из идеи Ламарка о внутреннем стремлении живых организмов к прогрессу; к представителям этой теории относится К. Бэр - основатель эмбриологии. Сальтационизм является модификацией телеогенеза и близок к теории катастрофизма. Суть его сводится к тому, что на начальном этапе зарождения жизни возник весь план будущего развития, а влияние внешних условий определяло лишь частные моменты эволюции. Эволюционные события происходят в результате скачкообразных изменений (сальтаций), прежде всего преобразований эмбриогенеза. Генетический антидарвинизм также имеет несколько течений: мутационизм, предадаптационизм, гибридогенез и др. В теории мутационизма мутационная изменчивость отождествлялась с эволюционными преобразованиями, что исключало необходимость процесса отбора как главной причины эволюции. Согласно теории предадаптационизма, приспособление возникает в результате не отбора, а единичной мутации, случайно оказавшейся полезной. Гибридогенез основан на представлении о постоянстве генов и утверждении, что комбинативная изменчивость - единственная причина эволюции.
Основные положения синтетической теории эволюции
Синтетическая теория эволюции — дальнейшее развитие дарвинизма. Она поддерживается большинством современных биологов [1, 3, 12, 14, 20, 26, 30, 33, 34, 38]. Эта теория включает анализ микро- и макроэволюции. Под микроэволюцией понимают совокупность эволюционных процессов, протекающих в популяциях и видах и приводящих к изменениям генофондов этих популяций и образованию новых видов. Считается, что микроэволюция проходит на основе мутационной изменчивости под контролем естественного отбора. Мутации служат единственным источником появления качественно новых признаков, а отбор - единственным творческим фактором микроэволюции, направляющим элементарные эволюционные изменения по пути формирования адаптации организмов к изменяющимся условиям внешней среды. На характер процессов микроэволюции оказывают влияние колебания численности популяций («волны жизни»), обмен генетической информацией между ними, их изоляция и дрейф генов. Микроэволюция ведет либо к изменению всего генофонда биологического вида как целого (филетическая эволюция), либо — при изоляции каких-либо популяций - к их обособлению от родительского вида в качестве новых форм (видообразование).
Под макроэволюцией понимают эволюционные преобразования, ведущие к формированию таксонов более высокого ранга, чем вид (родов, семейств, отрядов, классов и т.д.). Считают, что макроэволюция не имеет специфических механизмов и осуществляется только посредством процессов микроэволюции, будучи их интегрированным выражением. Накапливаясь, микроэволюционные процессы выражаются внешне в макроэволюционных явлениях, т.е. макроэволюция представляет собой обобщенную картину эволюционных изменений. Поэтому на уровне макроэволюции обнаруживаются общие тенденции, направления и закономерности эволюции живой природы, которые не поддаются наблюдению на уровне микроэволюции.
Основные положения синтетической теории эволюции обычно сводятся к четырем:
◊ главным фактором эволюции считается естественный отбор, интегрирующий и регулирующий действие всех остальных факторов (онтогенетической изменчивости, мутагенеза, гибридизации, миграции, изоляции, пульсации численности и др.);
◊ эволюция протекает дивергентно, постепенно, посредством отбора случайных мутаций, а новые формы образуются через наследственные изменения (сальтации, жизненность которых определяется отбором);
◊ эволюционные изменения случайны и ненаправленны; исходным материалом эволюции являются мутации; исходная организация популяции и изменения внешних условий ограничивают и канализируют наследственные изменения в направлении неограниченного прогресса;
◊ макроэволюция, ведущая к образованию надвидовых групп, осуществляется только посредством процессов микроэволюции, и каких-либо специфических механизмов возникновения новых форм жизни не существует.
В синтетической теории эволюции выделяют такие элементарные явления и факторы, как популяция - элементарная эволюционная структура; изменение генотипического состава популяции — элементарное эволюционное явление; генофонд популяции - элементарный эволюционный материал; мутационный процесс, «волны жизни», изоляция, естественный отбор - элементарные эволюционные факторы, и рассматривают движущий, дизруптивный и стабилизирующий отбор.
Движущий отбор благоприятствует лишь одному направлению изменчивости и не благоприятствует остальным ее вариантам. Под контролем движущего отбора генофонд популяции изменяется как целое и не происходит дивергенции дочерних форм. При этом в генофонде накапливаются и распространяются мутации, обеспечивающие изменение фенотипа в данном направлении. В популяции от поколения к поколению происходит изменение признака в определенном направлении.
Например, у некоторых видов животных возникает устойчивость к ядохимикатам. Широко известны случаи, когда у серых крыс и «вредных» насекомых вырабатывалась устойчивость к некоторым ядам. Это объясняется тем, что при воздействии ядов выживают особи, случайно оказавшиеся невосприимчивыми к этим ядам.
Дизруптивный отбор благоприятствует двум или нескольким направлениям изменчивости, но не благоприятствует среднему (промежуточному) состоянию признака. При действии такого отбора обычно возникает несколько отчетливо различающихся фенотипических форм (полиморфизм). Если разные направления дизруптивного отбора обусловлены различиями условий внешней среды в разных частях ареала данного вида, то населяющие их популяции приобретают устойчивые фенотипические и генотипические различия, имеющие важное приспособительное значение. При отсутствии возможности скрещивания между такими популяциями в результате изоляции друг от друга происходит их дальнейшая дивергенция вплоть до обособления в качестве новых видов.
Например, на океанических островах, где часты сильные ветры, насекомые с нормальными крыльями сдуваются в океан и погибают, а особи с повышенными аэродинамическими свойствами и с рудиментарными (остаточными, утратившими свое основное значение) крыльями сохраняются.
Стабилизирующий отбор благоприятствует сохранению в популяции оптимального в данных условиях фенотипа, который становится преобладающим. Этот отбор действует против проявления фенотипической изменчивости и наблюдается при длительном сохранении постоянных условий внешней среды. При длительном действии стабилизирующего отбора фенотипы некоторых видов организмов могут оставаться практически неизменными в течение миллионов лет.
Именно благодаря стабилизирующему отбору сохранились такие «живые ископаемые», как реликтовый таракан, мало изменившийся за последние 300-350 млн лет; кистеперая рыба латимерия, родственники которой были распространены в палеозое; голосеменное растение гинкго, появившееся в юрский период, и т.д. Другой пример - высокая устойчивость размеров и формы растений, опыляемых насекомыми: необходимо, чтобы цветки соответствовали строению и размерам тела насекомых-опылителей, в противном случае цветки не будут образовывать семян.
В конечном счете эволюцию можно представить как непрерывный процесс возникновения и развития новых адаптации (приспособлений) — совокупности физиологических, поведенческих, популяционных и других особенностей биологического вида, обеспечивающих возможность специфического образа жизни особей в определенных условиях внешней среды. Некоторые приспособления имеют частное значение, т.е. действуют в конкретных условиях среды; другие дают возможность выйти некоторой группе особей в новую адаптивную зону (тип местообитаний с определенной совокупностью экологических условий) и ведут к быстрому эволюционному развитию групп особей, к более высокому уровню организации.
§ 9.4. Развитие жизни на Земле
Криптозойский зон. Архейская и протерозойская эры
Остановимся на вопросе о том, как именно, по современным представлениям, развивалась жизнь на нашей планете [8, 14, 20, 25, 27].
В архейской эре возникли первые живые организмы. Они в качестве пищи использовали органические соединения «первичного бульона» (см. § 9.2). Важнейший этап эволюции жизни на Земле связан с возникновением фотосинтеза, впоследствии обусловившего разделение органического мира на растительный и животный. Цианеи - первые фотосинтезирующие организмы, прокариотические синезеленые «водоросли» и появившиеся затем эукариотические зеленые водоросли выделяли в атмосферу из океана свободный кислород, что способствовало возникновению бактерий, способных жить в аэробной среде. По-видимому, на границе архейской и протерозойской эр произошли еще два крупных эволюционных события: появились половой процесс и многоклеточность. Пути дальнейших эволюционных преобразований первых многоклеточных разошлись: одни перешли к неподвижному образу жизни и превратились в организмы типа губок; другие стали перемещаться по субстрату с помощью ресничек ~ от них произошли плоские черви; третьи сохранили плавающий образ жизни, приобрели рот, дали начало кишечнополостным.
В протерозойской эре в морях обитало много разнообразных водорослей, в том числе прикрепленных ко дну форм. Суша еще была безжизненной, но по берегам водоемов начались почвообразовательные процессы в результате деятельности бактерий и микроскопических водорослей. В протерозойских отложениях находят остатки представителей вполне сформировавшихся типов животных.
Фанерозойский зон. Палеозойская эра
В начале палеозойской эры растения населяют в основном моря; в ордовике-силуре появляются первые наземные растения, занимающие промежуточное положение между водорослями и наземными сосудистыми растениями. Так, псилофиты уже имели проводящую (сосудистую) систему, слабодифференцированные ткани, могли укрепляться в почве, но у них отсутствовали настоящие корни (как и настоящие побеги). Дальнейшая эволюция растений на суше шла в направлении расчленения тела на вегетативные органы и ткани, совершенствование сосудистой системы (обеспечивающей быстрое передвижение воды на большую высоту). В засушливом девоне широко распространяются хвощи, плауны, папоротникообразные. Еще большего развития достигает наземная растительность в каменноугольном периоде (карбоне),характеризующемся влажным и теплым климатом на протяжении всего года. Появляются голосеменные растения, произошедшие от семенных папоротников.
Переход к семенному размножению дал растениям много преимуществ, способствовавших их широкому расселению: зародыш в семенах был защищен от неблагоприятных условий оболочками и обеспечен пищей, у него стало диплоидное число хромосом, т.е. два сходных их набора. У части голосеменных (хвойных) процесс полового размножения уже не был связан с водой: опыление осуществлялось ветром, а семена имели приспособления для распространения животными. Крупные споровые растения вымерли в пермском периоде в связи со значительным иссушением и похолоданием климата.
Животный мир развивался чрезвычайно бурно и был представлен большим количеством разнообразных форм. Пышного расцвета достигла жизнь в кембрийских морях: губки, множество членистоногих, кораллов, иглокожие, моллюски, громадные членистоногие (хищные ракоскорпионы).
В геологических отложениях ордовика обнаружены остатки животных, имевших внутренний осевой скелет, - бесчелюстных позвоночных, отдаленными потомками которых являются современные миноги и миксины. Бесчелюстные питались организмами, обитавшими в илистом дне рек и озер, засасывая пищу ртом. У этих организмов (рис. 9.2) рот представлял собой просто отверстие, ведущее в пищеварительный тракт; передний отдел его был пронизан жаберными щелями, между которыми располагались опорные хрящевые жаберные дуги. В результате отбора совершенствовался аппарат для захвата живой добычи, гораздо более питательной по сравнению с илистым детритом: третья пара жаберных дуг превратилась в челюсти, усаженные зубами; жаберная мускулатура преобразовалась в челюстную и подъязычную. Так на основе существовавших структур - скелетных жаберных дуг, служивших опорой органов дыхания, возник ротовой аппарат хватательного типа. Новообразовавшиеся челюсти оказались очень стойким органом и сохранились в последующей эволюции позвоночных.
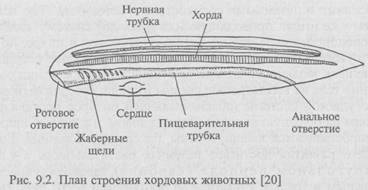
В силурийском периоде на сушу вместе с псилофитами вышли первые дышащие воздухом животные - членистоногие. В водоемах продолжалось бурное развитие низших позвоночных. Предполагается, что позвоночные возникли в мелких пресноводных водоемах и лишь затем переселились в моря.
В девоне позвоночные представлены тремя группами: двоякодышащими, лучеперыми и кистеперыми рыбами. В конце девона появились насекомые (кормовая база для будущих наземных позвоночных). Кистеперые рыбы были типично водными животными, но могли дышать атмосферным воздухом с помощью примитивных легких, представлявших собой выпячивания стенки кишки.
Поскольку уровень воды в реках и водоемах часто менялся, а многие водоемы полностью высыхали или промерзали зимой, что создавало неблагоприятную среду для рыб, то возникновение легких, т.е. приобретение способности дышать атмосферным воздухом, можно расценивать как своеобразную адаптацию к недостатку кислорода в воде. При пересыхании водоемов животные могли спастись, либо зарываясь в ил, либо мигрируя в поисках воды. По первому пути развивались двоякодышащие рыбы, строение которых почти не изменилось со времен девона и которые обитают и сейчас в мелких пересыхающих водоемах Африки. Лучеперые рыбы, имея плавники, поддерживающиеся отдельными костными лучами, широко распространились, и в настоящее время это самый большой по количеству видов класс позвоночных.
Приспособиться к жизни на суше смогли только кистеперые рыбы благодаря мускулистым конечностям и легким. В конце девона кистеперые рыбы дали начало первым земноводным - стегоцефалам.
На протяжении каменноугольного периода стегоцефалы жили, питались и размножались в воде. Они выползали на сушу, но не совершали сколько-нибудь значительных миграций. На суше у стегоцефалов не было врагов и имелся обильный корм - черви, членистоногие, достигавшие крупных размеров. Многие группы земноводных переходили к жизни на суше и возвращались в воду только для размножения. Среди стегоцефалов выделилась группа с хорошо развитыми конечностями и подвижной системой двух первых позвонков; они размножались в воде, но уходили по суше дальше амфибий, питались наземными животными, а затем и растениями. От них произошли рептилии и млекопитающие.
Пермский период отмечен поднятием суши, а также иссушением и похолоданием климата. Амфибии вымирали как из-за ухудшения климатических условий, так и вследствие истребления подвижными хищными рептилиями. Рептилии приобрели свойства, позволившие им порвать связь с водной средой: внутреннее оплодотворение и накопление желтка в яйцеклетке, что сделало возможным размножение на суше; ороговение кожи и более сложное строение почки, что способствовало резкому уменьшению потерь воды организмом; наличие грудной клетки, что обеспечило эффективный тип дыхания - всасывающий. Отсутствие конкуренции обусловило широкое распространение рептилий на суше и возвращение части их в водную среду.
Фанерозойский зон. Мезозойская и кайнозойская эры
Мезозойская эра характеризуется установлением теплого климата, близкого к современному тропическому. В триасе вымирают гигантские папоротники, древовидные хвощи, плауны, а расцвета достигают голосеменные растения.
В юрском периоде вымирают семенные папоротники и появляются первые покрытосеменные растения, постепенно распространившиеся на все материки вследствие ряда их преимуществ: покрытосеменные имеют развитую проводящую систему, цветок привлекает насекомых-опылителей, что обеспечивает надежность перекрестного опыления, зародыш снабжается запасами пищи и защищен оболочками и т.д. В животном мире достигают расцвета насекомые и рептилии; последние занимают господствующее положение и представлены большим количеством форм. В частности, в этот период появляются летающие ящеры, завоевавшие воздушную среду.
В меловом периоде специализация пресмыкающихся (рептилий) продолжается, они достигают громадных размеров; так масса некоторых динозавров превышала 50 т. Начинается параллельная эволюция цветковых растений и насекомых-опылителей. В конце мелового периода наступает похолодание, в результате чего сокращается ареал околоводной растительности; вымирают растительноядные, за ними хищные динозавры (крупные рептилии сохраняются лишь в тропическом поясе); в морях вымирают многие формы беспозвоночных и морские ящеры; наиболее приспособленными оказываются теплокровные животные - птицы и млекопитающие.
От представителей одного из отрядов рептилий произошли птицы. Полное разделение артериального и венозного кровотока обусловило их теплокровность. Они широко распространились по суше и дали начало множеству форм.
Появление млекопитающих связано с рядом крупных усложнений организации и функций организмов, возникших у представителей другого отряда пресмыкающихся. Млекопитающие возникли в триасе, но не могли конкурировать с хищными динозаврами и на протяжении 100 млн лет занимали подчиненное положение.
В начале кайнозойской эры установился теплый равномерный климат, а около 2 млн лет назад наступило оледенение значительной части Земли. Теплолюбивая растительность отступила на юг или вымерла, появилась холодоустойчивая травяная и кустарниковая растительность, на больших территориях леса сменились степью, полупустыней и пустыней, сформировались современные растительные сообщества.
Развитие животного мира характеризуется дальнейшей дифференциацией насекомых, интенсивным видообразованием у птиц и чрезвычайно быстрым прогрессивным развитием млекопитающих. При этом млекопитающие представлены тремя подклассами: однопроходными (утконос, ехидна), которые возникли независимо от других млекопитающих еще в юрском периоде от звероподобных рептилий; сумчатыми и плацентарными, которые произошли от общего предка в меловом периоде и сосуществовали до «взрыва» в эволюции плацентарных, в результате чего они вытеснили сумчатых с большинства континентов.
От наиболее примитивных — насекомоядных млекопитающих произошли первые хищные, давшие начало копытным, и приматы. В палеогене млекопитающие начинают завоевывать море (китообразные, ластоногие и др.), а к концу неогена встречаются уже все современные семейства млекопитающих. Одна из групп приматов (австралопитеки) стала родоначальницей ветви, ведущей к роду Человек.
Оледенения четвертичного периода, достигшие максимального распространения около 250 тыс. лет назад, привели к развитию холодоустойчивой фауны. На Северном Кавказе и в Крыму обитали мамонты, шерстистые носороги, северные олени, песцы, полярные куропатки. Образование больших масс льда вызвало понижение уровня Мирового океана, в результате чего обнажились материковые отмели Северной Америки и Северной Евразии и появились сухопутные «мосты», соединявшие Североамериканский континент с Евразийским (на месте нынешнего Берингова пролива), а также Британские острова с Европейским материком и т.д. По таким «мостам» происходил обмен видами, обусловивший формирование современной нам фауны материков. Изменения климата в четвертичном периоде кайнозойской эры оказали большое влияние на эволюцию предков человека.
§ 9.5. Человек: феномен, происхождение, биоэтика
Человек в системе животного мира
Человек — общественное существо, обладающее сознанием, разумом, субъект общественно-исторической деятельности и культуры. Человек познает и изменяет мир и самого себя, творит культуру и собственную историю. Сущность человека, его происхождение и назначение, место человека в мире были и остаются центральными проблемами философии, религии, науки и искусства.
Человек возник на Земле в ходе длительного и неравномерного эволюционного процесса — антропогенеза, многие этапы которого до конца не ясны [2, 21, 31, 38]. На планете Земля людям принадлежит уникальное место среди других существ, что обусловлено приобретением ими в процессе антропогенеза особого качества — социальной сущности. Это означает, что уже не биологические механизмы, а в первую очередь общественное устройство, производство, труд обеспечивают выживание, расселение, благополучие человечества. Однако социальность не противопоставляет людей остальной живой природе. Приобретение этого качества указывает лишь на то, что после некоторого момента развитие представителей вида Homo sapiens подчиняется законам не только биологического, но и в существенной степени общественного развития. Однако человек остается включенным в систему органического мира. Люди составляют своеобразный, но неотъемлемый компонент биосферы. Вследствие животного происхождения жизнедеятельность человеческого организма основывается на фундаментальных биологических механизмах, которые составляют его биологическое наследство.
Существует несколько точек зрения на соотношение социального и биологического в онтогенезе человека. Концепция панбиологизма исходит из того, что индивидуальное развитие человека целиком обусловлено генами, т.е. биологическими факторами. Согласно концепции пансоциологизма, все люди рождаются с одинаковыми биологическими задатками, а основную роль в развитии их способностей играет социальная среда. Еще одна концепция утверждает, что наследуются не сами способности как таковые, а лишь их задатки, в большей или меньшей степени проявляющиеся и реализующиеся в условиях социальной среды.
Общие принципы строения организма человека заложены в период возникновения первой клетки и формирования генетического кода. План строения тела человека такой же, как у всех представителей типа хордовых (см. рис. 9.2). Принадлежность человека к подтипу позвоночных определяется наличием осевого скелета в виде позвоночника, покровных тканей (черепа), защищающих переднюю часть нервной трубки, двух пар конечностей. Четырехкамерное сердце и редукция одной из двух дуг аорты, теплокровность, сильно развитая кора больших полушарий головного мозга, млечные железы, волосы на поверхности тела, дифференциация зубов на резцы, клыки и коренные свидетельствуют о принадлежности человека к классу млекопитающих. Зародыш человека развивается в теле матери и питается через плаценту, что определяет его положение в подклассе плацентарных. Такие признаки, как конечности хватательного типа (первый палец противопоставлен остальным), замена когтей плоскими ногтями, наличие одной пары млечных желез, у мужских особей семенники, спущенные в мошонку, хорошо развитые ключицы, замена молочных зубов на постоянные в ходе онтогенеза, наличие слепой кишки и др., позволяют отнести человека к отряду приматов. Эти признаки во многом предопределили ход эволюции отряда, приведшей к возникновению человека.
Характерные признаки человека как представителя отряда приматов хорошо объясняются приспособлением его к окружающей среде. Так, приспособления к древесному образу жизни у предка человека обусловили: способность его рук к тонким и разнообразным движениям, а также характер развития пищеварительной системы и снижение рождаемости. Последнее привело к расширению возможностей обучения потомства.
Особое значение имеет развитие у приматов специфического органа - мозга. Передвижение по деревьям в разных направлениях, на различные расстояния и с меняющейся скоростью, новой ориентировкой и новым прицелом перед прыжком определило высокое развитие ряда отделов мозга, особенно двигательных. Объем мозга в целом возрастает главным образом за счет коры больших полушарий, в которой увеличивается число борозд (извилин). Большая затылочная доля мозга связана с усложнением зрительных образов. При этом возрастает острота зрения, глазницы перемещаются с боковой поверхности на переднюю, благодаря чему поля зрения перекрываются и формируется бинокулярное зрение, которое дает стереоскопический эффект, позволяющий оценивать расстояния. Усложнение лобных долей объясняется необходимостью управлять мышечной деятельностью и голосовым аппаратом. Потребность в более совершенном различении звуков, нужном для звуковой сигнализации, привела к усложнению височных долей. Прогрессивно развивался мозжечок. Все эти изменения вызвали совершенно новое явление в эволюции мозга - доминирование (преобладание) коры больших полушарий над деятельностью низших мозговых центров. Увеличение мозга и преобразование органов чувств сопровождались ростом размеров лобной, теменной и затылочной костей черепа (рис. 9.3).
Такие признаки, как редукция хвостового отдела позвоночника, наличие аппендикса, большое количество извилин коры полушарий головного мозга, четыре основные группы крови, развитие мимической мускулатуры и др., позволяют отнести человека к подотряду человекообразных обезьян (антропоидов), или высших приматов. В этом подотряде выделяют надсемейство гоминид, которое включает человека и человекообразных обезьян. Наконец, семейство гоминид объединяет вид «человек разумный» и ископаемых людей (питекантропы, синантропы, неандертальцы и некоторые австралопитеки).
Особое значение для доказательства животного происхождения человека как результата длительной эволюции позвоночных имеет следующее обстоятельство. В эмбриональном периоде развития у зародыша человека закладываются двухкамерное сердце, шесть пар жаберных дуг, хвостовая артерия - признаки рыбообразных предков. От амфибий человек унаследовал плавательные перепонки между пальцами, которые имеются у зародыша. У новорожденных и детей до 5 лет наблюдается несовершенная терморегуляция, что указывает на происхождение от животных с непостоянной температурой тела. Головной мозг плода гладкий, без извилин, как у примитивных млекопитающих мезозойской эры. У шестинедельного зародыша имеется несколько пар млечных желез, закладывается также хвостовой отдел позвоночника, который затем редуцируется и превращается в копчик.
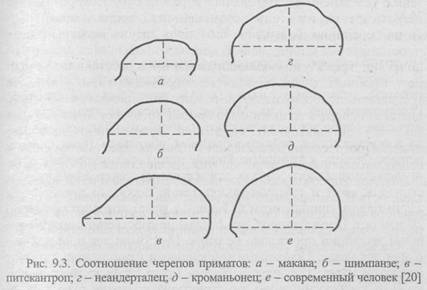
В XIX в. Φ. Мюллер и Э. Геккель сформулировали биогенетический закон, согласно которому онтогенез (индивидуальное развитие) каждой особи есть краткое и быстрое повторение филогенеза (исторического развития) вида, к которому эта особь относится. В соответствии с этим законом рассмотренные выше и многие другие признаки, возникающие у человека в эмбриональном периоде развития, могут быть интерпретированы как повторение признаков предков, причем эмбрионы всех позвоночных животных на ранних стадиях развития более сходны друг с другом, чем на более поздних.
Человек как вид имеет специфические, присущие только ему особенности: прямохождение, мощно развитую мускулатуру нижних конечностей, сводчатую стопу с сильно развитым первым пальцем, подвижную кисть руки, позвоночник с четырьмя изгибами, очень большой мозг по отношению к массе тела, крупные размеры мозгового и малые размеры лицевого отдела черепа, очень длительный период, предшествующий половому созреванию, членораздельную речь, общественный образ жизни. Эти особенности строения и физиологии человека - результат эволюции его животных предков.
Этапы антропогенеза
По современным представлениям, которые восходят к взглядам Ч. Дарвина, человек произошел от высокоразвитых обезьян. Эта гипотеза, называемая симиальной, имеет ряд модификаций. Рассмотрим одну из них. Многие независимые данные разных отраслей биологии подтверждают наибольшую близость человека к африканским человекообразным обезьянам (понгидам), в первую очередь к шимпанзе. Ранние этапы эволюции общего понгидно-гоминидного ствола представлены североафриканским египтопитеком и группой дриопитеков, особенно древнейшим африканским дриопитеком из Кении, возраст которого примерно 20 млн лет. Предполагают, что отделение собственно гоминидной ветви эволюции произошло не ранее 14—15 млн лет и не позднее 6 млн лет назад. Основными тенденциями гоминизации считают прямохождение, увеличение объема мозга, прежде всего новой коры, и дифференциацию его структуры, развитие руки как органа труда, удлинение периода роста и развития, освоение нового способа поведения - адаптацию к трудовой деятельности, что означало образование принципиально иной адаптационной ниши. Эволюция гоминид была неравномерной. По-видимому, скорость морфологических изменений не совпадала полностью с темпами биохимической эволюции; не было строгого соответствия прогресса морфофункциональной организации и культуры.
Примерно 5 млн лет назад гоминиды были представлены двуногими человекообезьянами - австралопитековыми, которые 4,5—1 млн лет назад довольно широко распространились в Африке, а возможно, и за ее пределами. В одной из периферийных популяций ранних австралопитековых могли сформироваться первые представители рода Homo, сосуществовавшие с австралопитековыми на большом временном промежутке, о чем свидетельствуют находки в Кении и Танзании. По другим представлениям, австралопитековые и Homo сформировались независимо при конвергентном развитии двуногого хождения в обеих линиях. Ископаемые остатки Homo (древностью около 2 млн лет) в ряде районов Восточной Африки (Танзания, Кения, Эфиопия) нередко находят вместе с «праорудиями» древнейшей каменной культуры человечества — олдовайской.
Принято считать, что первым был вид человек умелый (Homo habilis), сменившийся около 1,5 млн лет назад видом человек прямоходящий (Homo erectus). В дальнейшем происходило постепенное расширение экологической ниши Homo, и она поглотила экологические ниши австралопитековых. Примерно 1 млн лет назад Homo erectus стал единственным представителем гоминид на Земле. Этот вид обнаруживается в различных регионах Африки и Евразии примерно до 0,3 млн лет.
Человек современного типа (Homo sapiens) появился не позднее 40 тыс. лет назад. Его предками одни ученые считают ранних прогрессивных неандертальцев (Homo neandertalus), a другие — представителей конкурирующих ветвей Homo, линии которых разошлись всего 500 тыс. лет назад. Окончательное формирование современного типа человека датируют временем приблизительно 10—8 тыс. лет до н.э., когда он уже широко расселился по планете. Таким образом, развитие от первых человекообразных обезьян к современному человеку было очень сложным и не имело прямолинейного характера.
Существуют разногласия по вопросу о древности расовых различий, территории и количестве центров сапиентизации. Но большинство ученых полагает, что после расселения единый вид Homo sapiens распался на ряд рас - географических популяций. Они различаются по ряду внешних признаков (цвет кожи, форма головы, форма волос, форма носа, губ и т.п.), по частоте некоторых генов, детерминирующих биохимические признаки. Обычно выделяют три большие расы: экваториальную, или австрало-негроидную; евроазиатскую, или европеоидную; азиатско-американскую, или монголоидную, каждая из которых в свою очередь подразделяется на более мелкие расы.
С биологической точки зрения различия между расами не являются существенными и при смешении не препятствуют образованию полноценного плодовитого потомства. Общий уровень физического и умственного развития одинаков у всех людей. При этом признаки, лежащие в основе формирования расы, при переходе из одного географического района в другой изменяются постепенно, без резких скачков и часто перекрываются.
Соотношение природного и социального в антропогенезе было неодинаково на разных его этапах. В первобытном обществе гоминид прогресс социальной организации в значительной степени зависел от биологических изменений человека и в то же время процесс естественного отбора зависел от возникновения и развития общественных закономерностей и создания культурной среды. Относительная стабильность физического типа современного человека сохраняется примерно с середины позднего палеолита.
Считается, что в современном обществе отбор выступает как механизм поддержания сформировавшейся морфофункциональной организации в пределах видовой нормы реакции (стабилизирующий отбор) или как фактор внутривидового полиморфизма (дизруптивный отбор). Биологическое развитие современного человека проявляется обычно в виде разнонаправленных изменений различных признаков, а также структуры заболеваемости, темпов развития и т.д.
Биоэтика
В связи с развитием знаний о жизни у человека появилась возможность влиять на нее. Дело в том, что глубокое проникновение биологии в различные сферы общественной жизни потребовало усиления контроля со стороны общества за использованием ее научных достижений. Известно, что зачастую наиболее действенными средствами контроля оказываются общепринятые нормы морали. Эффективность норм нравственного регулирования объясняется силой привычки, традициями, обычаями, а также их универсальным характером. Моральные нормы чаще всего не нуждаются в контроле, так как они опираются на внутренний контроль, заложенный в душе каждого человека. Нормы морали очень динамичны, чутко реагируют на возникшие социальные проблемы. Они значительно опережают правовые нормы. Вероятно, это объясняет появление в последние годы новой сферы профессиональной этики - биоэтики (биологической или биомедицинской этики) [7].
Предметом изучения биоэтики являются быстро накапливающиеся достижения естествознания, в первую очередь биологии и медицины, глубокое их исследование и определение степени опасности в настоящем и будущем при их приложении к человеку и обществу в целом. Конечная цель биоэтики -разработка мер морального, а чаще законодательного характеpa, которые оградили бы каждого конкретного человека и человечество от нежелательных или губительных последствий внедрения новых и, к сожалению, не всегда до конца выверенных биологических и медицинских достижений.
Первоочередная проблема биоэтики связана с угрозой уничтожения всех форм жизни на Земле, ответственность за которую несут не только политики, но и ученые, особенно специалисты различных отраслей естествознания. Особое значение биоэтики обусловлено поразительными успехами современной генетики, внедрением искусственного оплодотворения, лекарственным «взрывом», наводнением внешней среды ксенобиотиками (чужеродными для организма соединениями — пестицидами, препаратами бытовой химии, лекарственными средствами и т.п.), разработкой новых способов регуляции деторождения, прогрессом трансплантологии, развитием вакцинопрофилактики инфекционных и вирусных заболеваний человека. В рамках биоэтики также привлекается внимание к проблемам гуманного отношения к животным, к совершенно новым и во многом еще не решенным проблемам ВИЧ-инфекции, принудительного лечения, врачебной тайны, эвтаназии (целесообразность поддержания жизни смертельно больного человека, допустимость использования человеком его «права на смерть»), к проведению научных экспериментов над животными и людьми, целесообразности применения достижений генетики для клонирования (копирования) животных и людей.
В процессе решения подобных и других биоэтических проблем утверждаются основные принципы биоэтики, среди которых чаще всего упоминают следующие:
◊ принцип единства жизни и этики, их глубокое соответствие и взаимообусловленность. Если жизнь - это высшее проявление упорядоченности, организованности в мире природы, то этика есть высшее выражение сил, противостоящих хаосу в обществе;
◊ принцип признания жизни в качестве высшей категории среди всех этических ценностей, принцип известного гуманиста А. Швейцера «преклонения перед жизнью»;
◊ принцип гармонизации системы человек - биосфера, выдвигающий в качестве актуальной задачи современности налаживание оптимальных взаимоотношений между человеком и природой и требующий от общества все более полного учета биологических оснований социального бытия, поиска путей превращения биосферы в ноосферу (сферу разума) и предотвращения возможности ее уничтожения.
Современная биоэтика основывается на принципе уважения к жизни и достоинству каждого человека, интересы которого во всех случаях должны ставиться выше интересов науки и общества. Одним из коренных положений биоэтики является автономия (неприкосновенность) психического и физического статуса человека. Заметим, что при изучении проблем биоэтики перекрещиваются интересы биологических, медицинских, философских и юридических наук. Поэтому успехи в научном и практическом решении проблем биоэтики в немалой степени зависят от налаживания плодотворного сотрудничества представителей различных областей знания.
ВОПРОСЫ ДЛЯ САМОКОНТРОЛЯ
- Что такое жизнь? Каковы основные свойства жизни?
- Какие основные уровни организации живой материи вы знаете?
- Как устроена и как функционирует клетка?
- Как, по современным представлениям, зародилась жизнь на Земле? Перечислите основные условия и факторы, способствовавшие этому.
- Какие основные принципы биологической эволюции вы знаете?
- Что такое микро- и макроэволюция? Как они соотносятся?
- В чем состоят основные позиции синтетической теории эволюции?
- В чем сходство и различия теории Ч. Дарвина и синтетической
теории эволюции?
- Когда зародилась жизнь и как происходило развитие жизни на ее самых ранних стадиях?
- Какими путями развивалась жизнь в архейском и протерозойском зонах? Назовите характерные формы жизни того времени.
- Как развивалась жизнь в палеозойскую эру? Какие формы жизни в это время вымирают, а какие появляются?
- В чем состоят основные изменения в живой природе в мезозойскую и кайнозойскую эры? Какие формы жизни были унаследованы от предыдущих эр, какие вымерли в эти эры, а какие появились?
- Как происходило становление человека как вида? Перечислите основные этапы антропогенеза.
- Каково соотношение биологического и социального в истории становления человечества? Продолжается ли в настоящее время биологическая эволюция человека?
- Что такое биоэтика, чем она занимается и каковы ее основные принципы?
ЛИТЕРАТУРА
- Айала Ф. Введение в популяционную и эволюционную генетику. М., 1984.
- Алексеев В.П. Становление человечества. М., 1984.
- Алихаян С.И., Акифьев А.П., Чернин Л.С. Общая генетика. М., 1985.
- Аллен Р.Д. Наука о жизни. М., 1981.
- Берг Л.С. Труды по теории эволюции. М., 1977.
- Биологический энциклопедический словарь. М., 1995.
- Биомедицинская этика / Под ред. В.И. Покровского. М., 1997.
- Бондаренко О.Б., Михайлова И.А. Палеонтология. Т. 1,2. М., 1997.
- Вайскопф В. Наука и удивительное. Как человек понимает природу. М., 1965.
- Вернадский В И. Живое вещество. М., 1978.
- Вилли К, Детъе В. Биология. М., 1975.
- Воронцов H.H. Теория эволюции: Истоки, постулаты и проблемы. М., 1984.
- Ганты Т. Жизнь и ее происхождение. М., 1984.
- Грант В. Эволюция организмов. М., 1980.
- Грин Н., Стаут У., Тейлор Д. Биология: В 3 т. М., 1993.
- Грядовой Д.И. Концепции современного естествознания: Структурный курс основ естествознания. М., 2000.
- Дарвин Ч. Происхождение видов путем естественного отбора. М., 1986.
- Иванов A.B. Происхождение многоклеточных животных. Л., 1968.
- Кемп П., Арме К. Введение в биологию. М., 1988.
- Мамонтов С.Г., Захаров В.Б., Козлова Т.А. Основы биологии. М., 1992.
- Матюшин Г.Н. У истоков человечества. М., 1982.
- Медников Б.М. Аксиомы биологии. М., 1982.
- Опарин А.И. Возникновение жизни на Земле. М., 1957.
- Опарин А.И. Материя, жизнь, интеллект. М., 1977.
- Орлов Ю.А. В мире древних животных. М., 1968.
- Северцов А.С. Основы теории эволюции. М., 1987.
- Татаринов Л.П. Палеонтология и эволюционное учение. М., 1985.
- Уровни организации биологических систем. М., 1980.
- Фолсом К.Э. Происхождение жизни. М., 1982.
- Фролов И.Т. Философия и история генетики. М., 1988.
- Хрисанова E.H., Перевозчиков И.В. Антропология. М., 2002.
- Шкловский И.С. Вселенная, жизнь, разум. М., 1980.
- Шмалъгаузен И.И. Вопросы дарвинизма. М., 1990.
- Шмалъгаузен И.И. Пути и закономерности эволюционного процесса. М., 1983.
- Шрёдингер Э. Что такое жизнь с точки зрения физики? М., 1972.
- Энгельгардт В.А. Познание явлений жизни. М,, 1984.
- Югай Г.А. Общая теория жизни. М., 1985.
- Яблоков A.B., Юсуфов А.Г. Эволюционное учение. М., 1998.
.
Ваш комментарий о книге
Обратно в раздел Наука
|
|